
|
|||||||
|

Fusion Protein:BRE-PRKCE |
Fusion Gene and Fusion Protein Summary |
 Fusion gene summary Fusion gene summary |
| Fusion partner gene information | Fusion gene name: BRE-PRKCE | FusionPDB ID: 10242 | FusionGDB2.0 ID: 10242 | Hgene | Tgene | Gene symbol | BRE | PRKCE | Gene ID | 9577 | 5581 |
| Gene name | BRISC and BRCA1 A complex member 2 | protein kinase C epsilon | |
| Synonyms | BRCC4|BRCC45|BRE | PKCE|nPKC-epsilon | |
| Cytomap | 2p23.2 | 2p21 | |
| Type of gene | protein-coding | protein-coding | |
| Description | BRISC and BRCA1-A complex member 2BRCA1-A complex subunit BREBRCA1/BRCA2-containing complex subunit 45BRCA1/BRCA2-containing complex, subunit 4brain and reproductive organ-expressed (TNFRSF1A modulator)brain and reproductive organ-expressed protein | protein kinase C epsilon type | |
| Modification date | 20200313 | 20200327 | |
| UniProtAcc | . | Q02156 Main function of 5'-partner protein: FUNCTION: Calcium-independent, phospholipid- and diacylglycerol (DAG)-dependent serine/threonine-protein kinase that plays essential roles in the regulation of multiple cellular processes linked to cytoskeletal proteins, such as cell adhesion, motility, migration and cell cycle, functions in neuron growth and ion channel regulation, and is involved in immune response, cancer cell invasion and regulation of apoptosis. Mediates cell adhesion to the extracellular matrix via integrin-dependent signaling, by mediating angiotensin-2-induced activation of integrin beta-1 (ITGB1) in cardiac fibroblasts. Phosphorylates MARCKS, which phosphorylates and activates PTK2/FAK, leading to the spread of cardiomyocytes. Involved in the control of the directional transport of ITGB1 in mesenchymal cells by phosphorylating vimentin (VIM), an intermediate filament (IF) protein. In epithelial cells, associates with and phosphorylates keratin-8 (KRT8), which induces targeting of desmoplakin at desmosomes and regulates cell-cell contact. Phosphorylates IQGAP1, which binds to CDC42, mediating epithelial cell-cell detachment prior to migration. In HeLa cells, contributes to hepatocyte growth factor (HGF)-induced cell migration, and in human corneal epithelial cells, plays a critical role in wound healing after activation by HGF. During cytokinesis, forms a complex with YWHAB, which is crucial for daughter cell separation, and facilitates abscission by a mechanism which may implicate the regulation of RHOA. In cardiac myocytes, regulates myofilament function and excitation coupling at the Z-lines, where it is indirectly associated with F-actin via interaction with COPB1. During endothelin-induced cardiomyocyte hypertrophy, mediates activation of PTK2/FAK, which is critical for cardiomyocyte survival and regulation of sarcomere length. Plays a role in the pathogenesis of dilated cardiomyopathy via persistent phosphorylation of troponin I (TNNI3). Involved in nerve growth factor (NFG)-induced neurite outgrowth and neuron morphological change independently of its kinase activity, by inhibition of RHOA pathway, activation of CDC42 and cytoskeletal rearrangement. May be involved in presynaptic facilitation by mediating phorbol ester-induced synaptic potentiation. Phosphorylates gamma-aminobutyric acid receptor subunit gamma-2 (GABRG2), which reduces the response of GABA receptors to ethanol and benzodiazepines and may mediate acute tolerance to the intoxicating effects of ethanol. Upon PMA treatment, phosphorylates the capsaicin- and heat-activated cation channel TRPV1, which is required for bradykinin-induced sensitization of the heat response in nociceptive neurons. Is able to form a complex with PDLIM5 and N-type calcium channel, and may enhance channel activities and potentiates fast synaptic transmission by phosphorylating the pore-forming alpha subunit CACNA1B (CaV2.2). In prostate cancer cells, interacts with and phosphorylates STAT3, which increases DNA-binding and transcriptional activity of STAT3 and seems to be essential for prostate cancer cell invasion. Downstream of TLR4, plays an important role in the lipopolysaccharide (LPS)-induced immune response by phosphorylating and activating TICAM2/TRAM, which in turn activates the transcription factor IRF3 and subsequent cytokines production. In differentiating erythroid progenitors, is regulated by EPO and controls the protection against the TNFSF10/TRAIL-mediated apoptosis, via BCL2. May be involved in the regulation of the insulin-induced phosphorylation and activation of AKT1. Phosphorylates NLRP5/MATER and may thereby modulate AKT pathway activation in cumulus cells (PubMed:19542546). {ECO:0000269|PubMed:11884385, ECO:0000269|PubMed:1374067, ECO:0000269|PubMed:15355962, ECO:0000269|PubMed:16757566, ECO:0000269|PubMed:17603037, ECO:0000269|PubMed:17875639, ECO:0000269|PubMed:17875724, ECO:0000269|PubMed:19542546}. | |
| Ensembl transtripts involved in fusion gene | ENST ids | ENST00000342045, ENST00000344773, ENST00000361704, ENST00000379624, ENST00000379632, ENST00000603461, | ENST00000394874, ENST00000467135, ENST00000306156, |
| Fusion gene scores for assessment (based on all fusion genes of FusionGDB 2.0) | * DoF score | 13 X 9 X 8=936 | 11 X 12 X 7=924 |
| # samples | 15 | 12 | |
| ** MAII score | log2(15/936*10)=-2.64154602908752 possibly effective Gene in Pan-Cancer Fusion Genes (peGinPCFGs). DoF>8 and MAII<0 | log2(12/924*10)=-2.94485844580754 possibly effective Gene in Pan-Cancer Fusion Genes (peGinPCFGs). DoF>8 and MAII<0 | |
| Fusion gene context | PubMed: BRE [Title/Abstract] AND PRKCE [Title/Abstract] AND fusion [Title/Abstract] | ||
| Fusion neoantigen context | PubMed: BRE [Title/Abstract] AND PRKCE [Title/Abstract] AND neoantigen [Title/Abstract] | ||
| Most frequent breakpoint (based on all fusion genes of FusionGDB 2.0) | BRE(28268666)-PRKCE(46313347), # samples:3 | ||
| Anticipated loss of major functional domain due to fusion event. | BRE-PRKCE seems lost the major protein functional domain in Hgene partner, which is a CGC by not retaining the major functional domain in the partially deleted in-frame ORF. BRE-PRKCE seems lost the major protein functional domain in Hgene partner, which is a CGC by not retaining the major functional domain in the partially deleted in-frame ORF. BRE-PRKCE seems lost the major protein functional domain in Hgene partner, which is a essential gene by not retaining the major functional domain in the partially deleted in-frame ORF. BRE-PRKCE seems lost the major protein functional domain in Hgene partner, which is a essential gene by not retaining the major functional domain in the partially deleted in-frame ORF. | ||
| * DoF score (Degree of Frequency) = # partners X # break points X # cancer types ** MAII score (Major Active Isofusion Index) = log2(# samples/DoF score*10) |
| Gene ontology of each fusion partner gene with evidence of Inferred from Direct Assay (IDA) from Entrez |
| Partner | Gene | GO ID | GO term | PubMed ID |
| Hgene | BRE | GO:0043066 | negative regulation of apoptotic process | 15465831 |
| Tgene | PRKCE | GO:0006468 | protein phosphorylation | 18556656 |
| Tgene | PRKCE | GO:0018105 | peptidyl-serine phosphorylation | 15695813 |
| Four levels of functional features of fusion genes Go to FGviewer search page for the most frequent breakpoint (https://ccsmweb.uth.edu/FGviewer/chr2:28268666/chr2:46313347) - FGviewer provides the online visualization of the retention search of the protein functional features across DNA, RNA, protein, and pathological levels. - How to search 1. Put your fusion gene symbol. 2. Press the tab key until there will be shown the breakpoint information filled. 4. Go down and press 'Search' tab twice. 4. Go down to have the hyperlink of the search result. 5. Click the hyperlink. 6. See the FGviewer result for your fusion gene. |
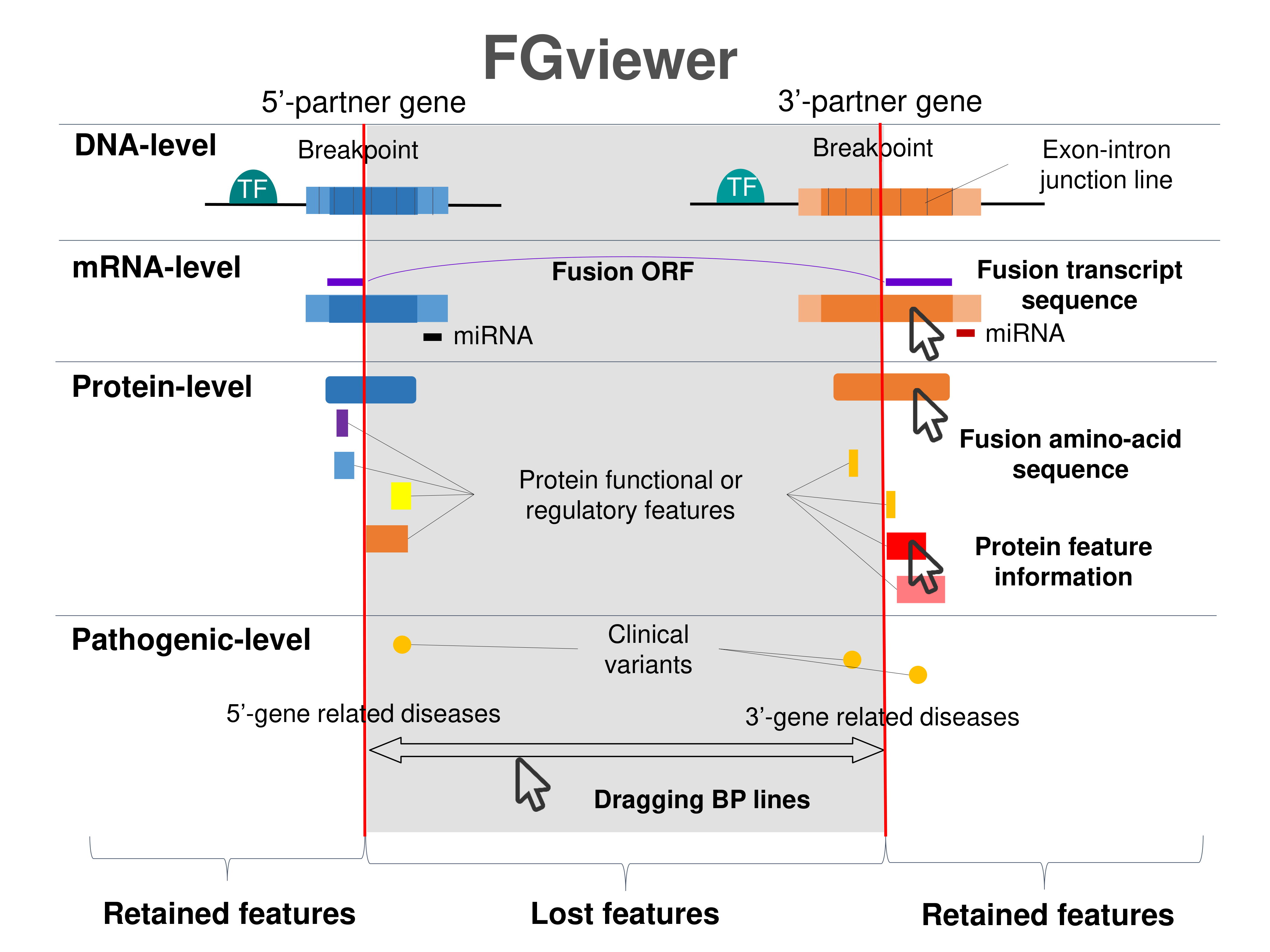 |
| Retention analysis results of each fusion partner protein across 39 protein features of UniProt such as six molecule processing features, 13 region features, four site features, six amino acid modification features, two natural variation features, five experimental info features, and 3 secondary structure features, are available here. |
| Fusion gene breakpoints across BRE (5'-gene) * Click on the image to open the UCSC genome browser with custom track showing this image in a new window. |
 |
| Fusion gene breakpoints across PRKCE (3'-gene) * Click on the image to open the UCSC genome browser with custom track showing this image in a new window. |
 |
Top |
Fusion Amino Acid Sequences |
| Fusion information from ORFfinder translation from full-length transcript sequence from FusionPDB. |
| Henst | Tenst | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand | Seq length (transcript) | BP loci (transcript) | Predicted start (transcript) | Predicted stop (transcript) | Seq length (amino acids) |
| ENST00000344773 | BRE | chr2 | 28268666 | + | ENST00000306156 | PRKCE | chr2 | 46313347 | + | 4594 | 708 | 138 | 1484 | 448 |
| ENST00000379624 | BRE | chr2 | 28268666 | + | ENST00000306156 | PRKCE | chr2 | 46313347 | + | 4573 | 687 | 117 | 1463 | 448 |
| ENST00000342045 | BRE | chr2 | 28268666 | + | ENST00000306156 | PRKCE | chr2 | 46313347 | + | 4597 | 711 | 141 | 1487 | 448 |
| ENST00000379632 | BRE | chr2 | 28268666 | + | ENST00000306156 | PRKCE | chr2 | 46313347 | + | 4596 | 710 | 140 | 1486 | 448 |
| ENST00000361704 | BRE | chr2 | 28268666 | + | ENST00000306156 | PRKCE | chr2 | 46313347 | + | 4530 | 644 | 74 | 1420 | 448 |
| DeepORF prediction of the coding potential based on the fusion transcript sequence of in-frame fusion genes. DeepORF is a coding potential classifier based on convolutional neural network by comparing the real Ribo-seq data. If the no-coding score < 0.5 and coding score > 0.5, then the in-frame fusion transcript is predicted as being likely translated. |
| Henst | Tenst | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand | No-coding score | Coding score |
| ENST00000344773 | ENST00000306156 | BRE | chr2 | 28268666 | + | PRKCE | chr2 | 46313347 | + | 0.000223038 | 0.999777 |
| ENST00000379624 | ENST00000306156 | BRE | chr2 | 28268666 | + | PRKCE | chr2 | 46313347 | + | 0.000216316 | 0.9997837 |
| ENST00000342045 | ENST00000306156 | BRE | chr2 | 28268666 | + | PRKCE | chr2 | 46313347 | + | 0.000207263 | 0.99979275 |
| ENST00000379632 | ENST00000306156 | BRE | chr2 | 28268666 | + | PRKCE | chr2 | 46313347 | + | 0.000207236 | 0.99979275 |
| ENST00000361704 | ENST00000306156 | BRE | chr2 | 28268666 | + | PRKCE | chr2 | 46313347 | + | 0.000218069 | 0.9997819 |
| Predicted full-length fusion amino acid sequences. For individual full-length fusion transcript sequence from FusionPDB, we ran ORFfinder and chose the longest ORF among all the predicted ones. |
Get the fusion protein sequences from here. |
| Fusion protein sequence information is available in the fasta format. >FusionGDB ID_FusionGDB isoform ID_FGname_Hgene_Hchr_Hbp_Henst_Tgene_Tchr_Tbp_Tenst_length(fusion AA) seq_BP |
Top |
Fusion Protein Breakpoint Sequences for BRE-PRKCE |
| +/-13 AA sequence from the breakpoints of the fusion protein sequences. |
| Hgene | Hchr | Hbp | Tgene | Tchr | Tbp | Length(fusion protein) | BP in fusion protein | Peptide |
| BRE | chr2 | 28268666 | PRKCE | chr2 | 46313347 | 644 | 190 | PVDFSNIPTYLLKDRLFFVMEYVNGG |
| BRE | chr2 | 28268666 | PRKCE | chr2 | 46313347 | 687 | 190 | PVDFSNIPTYLLKDRLFFVMEYVNGG |
| BRE | chr2 | 28268666 | PRKCE | chr2 | 46313347 | 708 | 190 | PVDFSNIPTYLLKDRLFFVMEYVNGG |
| BRE | chr2 | 28268666 | PRKCE | chr2 | 46313347 | 710 | 190 | PVDFSNIPTYLLKDRLFFVMEYVNGG |
| BRE | chr2 | 28268666 | PRKCE | chr2 | 46313347 | 711 | 190 | PVDFSNIPTYLLKDRLFFVMEYVNGG |
Top |
Potential FusionNeoAntigen Information of BRE-PRKCE in HLA I |
| Multiple sequence alignments of the potential FusionNeoAntigens per fusion breakpoints. If the MSA is empty, then it means that there were predicted fusion neoantigens in this fusion breakpoint, but those predicted fusion neoantigens were not across the breakpoint, which is not fusion-specific. |
| BRE-PRKCE_28268666_46313347.msa |
| Potential FusionNeoAntigen Information * We used NetMHCpan v4.1 (%rank<0.5) and deepHLApan v1.1 (immunogenic score>0.5) |
| Fusion gene | Hchr | Hbp | Tgene | Tchr | Tbp | HLA I | FusionNeoAntigen peptide | Binding score | Immunogenic score | Neoantigen start (at BP 13) | Neoantigen end (at BP 13) |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:22 | LLKDRLFFV | 0.996 | 0.5113 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:13 | LLKDRLFFV | 0.9904 | 0.5255 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:60 | LLKDRLFFV | 0.9771 | 0.5644 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:24 | LLKDRLFFV | 0.9768 | 0.5808 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:30 | LLKDRLFFV | 0.9768 | 0.5808 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:67 | LLKDRLFFV | 0.9768 | 0.5808 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:11 | LLKDRLFFV | 0.9756 | 0.5974 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:21 | LLKDRLFFV | 0.9734 | 0.7025 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:35 | LLKDRLFFV | 0.9339 | 0.6264 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:04 | LLKDRLFFV | 0.9246 | 0.5621 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A32:13 | YLLKDRLFF | 0.8931 | 0.8399 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:25 | YLLKDRLFF | 0.8729 | 0.8226 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:29 | LLKDRLFFV | 0.8629 | 0.5859 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:20 | LLKDRLFFV | 0.843 | 0.5852 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:02 | YLLKDRLFF | 0.8214 | 0.841 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B13:01 | YLLKDRLFF | 0.0205 | 0.8557 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:24 | YLLKDRLFFV | 0.9963 | 0.5504 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:30 | YLLKDRLFFV | 0.9963 | 0.5504 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:60 | YLLKDRLFFV | 0.9963 | 0.5341 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:67 | YLLKDRLFFV | 0.9963 | 0.5504 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:11 | YLLKDRLFFV | 0.9963 | 0.5646 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:13 | YLLKDRLFFV | 0.9944 | 0.5114 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:21 | YLLKDRLFFV | 0.9937 | 0.6721 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:05 | KDRLFFVMEY | 0.9855 | 0.8617 | 12 | 22 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:04 | YLLKDRLFFV | 0.9831 | 0.5846 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:02 | KDRLFFVMEY | 0.9807 | 0.5429 | 12 | 22 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:04 | KDRLFFVMEY | 0.9789 | 0.6088 | 12 | 22 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:35 | YLLKDRLFFV | 0.9727 | 0.5928 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:29 | YLLKDRLFFV | 0.8711 | 0.5549 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:20 | YLLKDRLFFV | 0.8515 | 0.5532 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:05 | LLKDRLFFV | 0.995 | 0.5235 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:07 | LLKDRLFFV | 0.9773 | 0.676 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:01 | LLKDRLFFV | 0.9768 | 0.5808 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:05 | YLLKDRLFF | 0.4334 | 0.7693 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:67 | TYLLKDRLF | 0.3328 | 0.8982 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:80 | TYLLKDRLF | 0.3328 | 0.8982 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:46 | TYLLKDRLF | 0.3126 | 0.7935 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:10 | TYLLKDRLF | 0.3071 | 0.9321 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:07 | YLLKDRLFFV | 0.9964 | 0.6324 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:01 | YLLKDRLFFV | 0.9963 | 0.5504 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:03 | KDRLFFVMEY | 0.7919 | 0.8836 | 12 | 22 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:03 | LLKDRLFFV | 0.9982 | 0.6526 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:14 | LLKDRLFFV | 0.9758 | 0.6339 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:06 | LLKDRLFFV | 0.9734 | 0.7025 | 10 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A32:01 | YLLKDRLFF | 0.8882 | 0.9055 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:39 | YLLKDRLFF | 0.8703 | 0.667 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:27 | YLLKDRLFF | 0.8663 | 0.8322 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:24 | YLLKDRLFF | 0.8487 | 0.8336 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B15:20 | YLLKDRLFF | 0.4411 | 0.8449 | 9 | 18 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:17 | TYLLKDRLF | 0.4152 | 0.9334 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C07:02 | TYLLKDRLF | 0.3328 | 0.8982 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C14:03 | TYLLKDRLF | 0.0116 | 0.8995 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C14:02 | TYLLKDRLF | 0.0116 | 0.8995 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-C06:06 | TYLLKDRLF | 0.0109 | 0.961 | 8 | 17 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:03 | YLLKDRLFFV | 0.9977 | 0.6185 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:06 | YLLKDRLFFV | 0.9937 | 0.6721 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-A02:14 | YLLKDRLFFV | 0.9934 | 0.6221 | 9 | 19 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:08 | KDRLFFVMEY | 0.9808 | 0.7168 | 12 | 22 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | HLA-B27:10 | KDRLFFVMEY | 0.9792 | 0.7877 | 12 | 22 |
Top |
Potential FusionNeoAntigen Information of BRE-PRKCE in HLA II |
| Multiple sequence alignments of the potential FusionNeoAntigens per fusion breakpoints. If the MSA is empty, then it means that there were predicted fusion neoantigens in this fusion breakpoint, but those predicted fusion neoantigens were not across the breakpoint, which is not fusion-specific. |
| BRE-PRKCE_28268666_46313347.msa |
| Potential FusionNeoAntigen Information * We used NetMHCIIpan v4.1 (%rank<0.5). |
| Fusion gene | Hchr | Hbp | Tgene | Tchr | Tbp | HLA II | FusionNeoAntigen peptide | Neoantigen start (at BP 13) | Neoantigen end (at BP 13) |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0109 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0113 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0115 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0469 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0482 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0701 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0703 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0704 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0705 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0706 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0707 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0708 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0709 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0711 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0712 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0713 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0714 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0715 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0716 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0717 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0719 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0807 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0819 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0825 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-0834 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1216 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1222 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1446 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1448 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1502 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1508 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1511 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1514 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1515 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1519 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1526 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1527 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1529 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1530 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1531 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1534 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1538 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1539 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1544 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1547 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1601 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1602 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1603 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1604 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1605 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1607 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1608 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1609 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1610 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1611 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1612 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1614 | PVDFSNIPTYLLKDR | 0 | 15 |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 | DRB1-1616 | PVDFSNIPTYLLKDR | 0 | 15 |
Top |
Fusion breakpoint peptide structures of BRE-PRKCE |
| 3D structures of the fusion breakpoint peptide of 14AA sequence that have potential fusion neoantigens * The minimum length of the amino acid sequence in RoseTTAFold is 14AA. Here, we predicted the 14AA fusion protein breakpoint sequence not the fusion neoantigen peptide, which is shorter than 14 AA. |
| File name | BPseq | Hgene | Tgene | Hchr | Hbp | Tchr | Tbp | AAlen |
| 3898 | IPTYLLKDRLFFVM | BRE | PRKCE | chr2 | 28268666 | chr2 | 46313347 | 711 |
Top |
Filtering FusionNeoAntigens Through Checking the Interaction with HLAs in 3D of BRE-PRKCE |
| Virtual screening between 25 HLAs (from PDB) and FusionNeoAntigens * We used Glide to predict the interaction between HLAs and neoantigens. |
| HLA allele | PDB ID | File name | BPseq | Docking score | Glide score |
| HLA-B14:02 | 3BVN | 3898 | IPTYLLKDRLFFVM | -6.35143 | -7.38673 |
| HLA-B14:02 | 3BVN | 3898 | IPTYLLKDRLFFVM | -5.20162 | -5.31502 |
| HLA-B52:01 | 3W39 | 3898 | IPTYLLKDRLFFVM | -6.03049 | -6.14389 |
| HLA-B52:01 | 3W39 | 3898 | IPTYLLKDRLFFVM | -5.12323 | -6.15853 |
| HLA-A24:02 | 5HGA | 3898 | IPTYLLKDRLFFVM | -9.81198 | -9.92538 |
| HLA-A24:02 | 5HGA | 3898 | IPTYLLKDRLFFVM | -7.16433 | -8.19963 |
| HLA-B44:05 | 3DX8 | 3898 | IPTYLLKDRLFFVM | -6.46131 | -6.57471 |
| HLA-B44:05 | 3DX8 | 3898 | IPTYLLKDRLFFVM | -3.08576 | -4.12106 |
| HLA-A02:01 | 6TDR | 3898 | IPTYLLKDRLFFVM | -5.03933 | -5.15273 |
Top |
Vaccine Design for the FusionNeoAntigens of BRE-PRKCE |
| mRNA and peptide sequences of FusionNeoAntigens that have potential interaction with HLA-Is. |
| Fusion gene | Hchr | Hbp | Tchr | Tbp | Start in +/-13AA | End in +/-13AA | FusionNeoAntigen peptide sequence | FusionNeoAntigen RNA sequence |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 10 | 19 | LLKDRLFFV | CTTCTCAAGGACCGCCTCTTTTTCGTC |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 12 | 22 | KDRLFFVMEY | AAGGACCGCCTCTTTTTCGTCATGGAATAT |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 8 | 17 | TYLLKDRLF | ACATACCTTCTCAAGGACCGCCTCTTT |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 9 | 18 | YLLKDRLFF | TACCTTCTCAAGGACCGCCTCTTTTTC |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 9 | 19 | YLLKDRLFFV | TACCTTCTCAAGGACCGCCTCTTTTTCGTC |
| mRNA and peptide sequences of FusionNeoAntigens that have potential interaction with HLA-IIs. |
| Fusion gene | Hchr | Hbp | Tchr | Tbp | Start in +/-13AA | End in +/-13AA | FusionNeoAntigen peptide | FusionNEoAntigen RNA sequence |
| BRE-PRKCE | chr2 | 28268666 | chr2 | 46313347 | 0 | 15 | PVDFSNIPTYLLKDR | CCCGTAGATTTCAGCAATATCCCCACATACCTTCTCAAGGACCGC |
Top |
Information of the samples that have these potential fusion neoantigens of BRE-PRKCE |
| These samples were reported as having these fusion breakpoints. For individual breakpoints, we checked the open reading frames considering multiple gene isoforms and chose the in-frame fusion genes only. Then, we made fusion protein sequences and predicted the fusion neoantigens. These fusion-positive samples may have these potential fusion neoantigens. |
| Cancer type | Fusion gene | Hchr | Hbp | Henst | Tchr | Tbp | Tenst | Sample |
| BRCA | BRE-PRKCE | chr2 | 28268666 | ENST00000342045 | chr2 | 46313347 | ENST00000306156 | TCGA-C8-A134-01A |
Top |
Potential target of CAR-T therapy development for BRE-PRKCE |
| Predicted 3D structure. We used RoseTTAFold. |
| Retention analysis result of each fusion partner protein across 39 protein features of UniProt such as six molecule processing features, 13 region features, four site features, six amino acid modification features, two natural variation features, five experimental info features, and 3 secondary structure features. Here, to provide the retention of the transmembrane domain, we only show the protein feature retention information of those transmembrane features * Minus value of BPloci means that the break point is located before the CDS. |
| - In-frame and retained 'Transmembrane'. |
| Partner | Gene | Hbp | Tbp | ENST | Strand | BPexon | TotalExon | Protein feature loci | *BPloci | TotalLen | Protein feature | Protein feature note |
| Subcellular localization prediction of the transmembrane domain retained fusion proteins * We used DeepLoc 1.0. The order of the X-axis of the barplot is as follows: Entry_ID, Localization, Type, Nucleus, Cytoplasm, Extracellular, Mitochondrion, Cell_membrane, Endoplasmic_reticulum, Plastid, Golgi.apparatus, Lysosome.Vacuole, Peroxisome. Y-axis is the output score of DeepLoc. Clicking the image will open a new tab with a large image. |
| Hgene | Hchr | Hbp | Henst | Tgene | Tchr | Tbp | Tenst | DeepLoc result |
Top |
Related Drugs to BRE-PRKCE |
| Drugs used for this fusion-positive patient. (Manual curation of PubMed, 04-30-2022 + MyCancerGenome) |
| Hgene | Tgene | Drug | Source | PMID |
Top |
Related Diseases to BRE-PRKCE |
| Diseases that have this fusion gene. (Manual curation of PubMed, 04-30-2022 + MyCancerGenome) |
| Hgene | Tgene | Disease | Source | PMID |
| Diseases associated with fusion partners. (DisGeNet 4.0) |
| Partner | Gene | Disease ID | Disease name | # pubmeds | Source |
| Tgene | PRKCE | C0020429 | Hyperalgesia | 3 | CTD_human |
| Tgene | PRKCE | C0458247 | Allodynia | 3 | CTD_human |
| Tgene | PRKCE | C0751211 | Hyperalgesia, Primary | 3 | CTD_human |
| Tgene | PRKCE | C0751212 | Hyperalgesia, Secondary | 3 | CTD_human |
| Tgene | PRKCE | C0751213 | Tactile Allodynia | 3 | CTD_human |
| Tgene | PRKCE | C0751214 | Hyperalgesia, Thermal | 3 | CTD_human |
| Tgene | PRKCE | C2936719 | Mechanical Allodynia | 3 | CTD_human |
| Tgene | PRKCE | C0002152 | Alloxan Diabetes | 1 | CTD_human |
| Tgene | PRKCE | C0009402 | Colorectal Carcinoma | 1 | CTD_human;UNIPROT |
| Tgene | PRKCE | C0009404 | Colorectal Neoplasms | 1 | CTD_human |
| Tgene | PRKCE | C0011853 | Diabetes Mellitus, Experimental | 1 | CTD_human |
| Tgene | PRKCE | C0011881 | Diabetic Nephropathy | 1 | CTD_human |
| Tgene | PRKCE | C0013146 | Drug abuse | 1 | CTD_human |
| Tgene | PRKCE | C0013170 | Drug habituation | 1 | CTD_human |
| Tgene | PRKCE | C0013222 | Drug Use Disorders | 1 | CTD_human |
| Tgene | PRKCE | C0017667 | Nodular glomerulosclerosis | 1 | CTD_human |
| Tgene | PRKCE | C0023903 | Liver neoplasms | 1 | CTD_human |
| Tgene | PRKCE | C0027051 | Myocardial Infarction | 1 | CTD_human |
| Tgene | PRKCE | C0029231 | Organic Mental Disorders, Substance-Induced | 1 | CTD_human |
| Tgene | PRKCE | C0033141 | Cardiomyopathies, Primary | 1 | CTD_human |
| Tgene | PRKCE | C0036529 | Myocardial Diseases, Secondary | 1 | CTD_human |
| Tgene | PRKCE | C0038433 | Streptozotocin Diabetes | 1 | CTD_human |
| Tgene | PRKCE | C0038580 | Substance Dependence | 1 | CTD_human |
| Tgene | PRKCE | C0038586 | Substance Use Disorders | 1 | CTD_human |
| Tgene | PRKCE | C0151744 | Myocardial Ischemia | 1 | CTD_human |
| Tgene | PRKCE | C0236969 | Substance-Related Disorders | 1 | CTD_human |
| Tgene | PRKCE | C0242231 | Coronary Stenosis | 1 | CTD_human |
| Tgene | PRKCE | C0345904 | Malignant neoplasm of liver | 1 | CTD_human |
| Tgene | PRKCE | C0400966 | Non-alcoholic Fatty Liver Disease | 1 | CTD_human |
| Tgene | PRKCE | C0740858 | Substance abuse problem | 1 | CTD_human |
| Tgene | PRKCE | C0878544 | Cardiomyopathies | 1 | CTD_human |
| Tgene | PRKCE | C1510472 | Drug Dependence | 1 | CTD_human |
| Tgene | PRKCE | C3241937 | Nonalcoholic Steatohepatitis | 1 | CTD_human |
| Tgene | PRKCE | C4316881 | Prescription Drug Abuse | 1 | CTD_human |
| Tgene | PRKCE | C4721453 | Peripheral Nervous System Diseases | 1 | CTD_human |
 Copyright 2023-Present
Copyright 2023-PresentThe University of Texas Health Science Center at Houston (UTHealth)
Web File Viewing | Emergency Information
Campus Carry|Site Policies