| UTHEALTH HOME ABOUT SBMI A-Z WEBMAIL INSIDE THE UNIVERSITY |

|
|||||||
|

Fusion Protein:ASXL2-HADHA |
Fusion Protein Summary |
 Fusion gene summary Fusion gene summary |
| Fusion partner gene information | Fusion gene name: ASXL2-HADHA | FusionPDB ID: 7307 | FusionGDB2.0 ID: 7307 | Hgene | Tgene | Gene symbol | ASXL2 | HADHA | Gene ID | 55252 | 3030 |
| Gene name | ASXL transcriptional regulator 2 | hydroxyacyl-CoA dehydrogenase trifunctional multienzyme complex subunit alpha | |
| Synonyms | ASXH2|SHAPNS | ECHA|GBP|HADH|LCEH|LCHAD|MTPA|TP-ALPHA | |
| Cytomap | 2p23.3 | 2p23.3 | |
| Type of gene | protein-coding | protein-coding | |
| Description | putative Polycomb group protein ASXL2additional sex combs like 2, transcriptional regulatoradditional sex combs-like protein 2polycomb group protein ASXH2 | trifunctional enzyme subunit alpha, mitochondrial3-ketoacyl-Coenzyme A (CoA) thiolase, alpha subunit3-oxoacyl-CoA thiolase78 kDa gastrin-binding proteingastrin-binding proteinhydroxyacyl-CoA dehydrogenase/3-ketoacyl-CoA thiolase/enoyl-CoA hydratase ( | |
| Modification date | 20200313 | 20200313 | |
| UniProtAcc | Q76L83 | P40939 | |
| Ensembl transtripts involved in fusion gene | ENST ids | ENST00000336112, ENST00000435504, ENST00000272341, ENST00000497092, ENST00000404843, | ENST00000457468, ENST00000461025, ENST00000380649, |
| Fusion gene scores for assessment (based on all fusion genes of FusionGDB 2.0) | * DoF score | 18 X 11 X 10=1980 | 7 X 6 X 7=294 |
| # samples | 24 | 9 | |
| ** MAII score | log2(24/1980*10)=-3.04439411935845 possibly effective Gene in Pan-Cancer Fusion Genes (peGinPCFGs). DoF>8 and MAII<0 | log2(9/294*10)=-1.70781924850669 possibly effective Gene in Pan-Cancer Fusion Genes (peGinPCFGs). DoF>8 and MAII<0 | |
| Context (manual curation of fusion genes in FusionPDB) | PubMed: ASXL2 [Title/Abstract] AND HADHA [Title/Abstract] AND fusion [Title/Abstract] | ||
| Most frequent breakpoint (based on all fusion genes of FusionGDB 2.0) | ASXL2(26101035)-HADHA(26432758), # samples:2 | ||
| Anticipated loss of major functional domain due to fusion event. | ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a CGC by not retaining the major functional domain in the partially deleted in-frame ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a CGC by not retaining the major functional domain in the partially deleted in-frame ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a essential gene by not retaining the major functional domain in the partially deleted in-frame ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a essential gene by not retaining the major functional domain in the partially deleted in-frame ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a CGC due to the frame-shifted ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a epigenetic factor due to the frame-shifted ORF. ASXL2-HADHA seems lost the major protein functional domain in Hgene partner, which is a essential gene due to the frame-shifted ORF. ASXL2-HADHA seems lost the major protein functional domain in Tgene partner, which is a cell metabolism gene due to the frame-shifted ORF. | ||
| * DoF score (Degree of Frequency) = # partners X # break points X # cancer types ** MAII score (Major Active Isofusion Index) = log2(# samples/DoF score*10) |
| Gene ontology of each fusion partner gene with evidence of Inferred from Direct Assay (IDA) from Entrez |
| Partner | Gene | GO ID | GO term | PubMed ID |
| Hgene | ASXL2 | GO:0035360 | positive regulation of peroxisome proliferator activated receptor signaling pathway | 21047783 |
| Hgene | ASXL2 | GO:0045600 | positive regulation of fat cell differentiation | 21047783 |
| Hgene | ASXL2 | GO:0045944 | positive regulation of transcription by RNA polymerase II | 21047783 |
| Tgene | HADHA | GO:0035965 | cardiolipin acyl-chain remodeling | 23152787 |
| Fusion gene breakpoints across ASXL2 (5'-gene) * Click on the image to open the UCSC genome browser with custom track showing this image in a new window. |
 |
| Fusion gene breakpoints across HADHA (3'-gene) * Click on the image to open the UCSC genome browser with custom track showing this image in a new window. |
 |
Top |
Fusion Gene Sample Information |
| Fusion gene information from FusionGDB2.0. |
| Fusion gene information from two resources (ChiTars 5.0 and ChimerDB 4.0) * All genome coordinats were lifted-over on hg19. * Click on the break point to see the gene structure around the break point region using the UCSC Genome Browser. |
| Source | Disease | Sample | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand |
| ChimerDB4 | ESCA | TCGA-L5-A43M-01A | ASXL2 | chr2 | 26101035 | - | HADHA | chr2 | 26432758 | - |
| ChimerDB4 | ESCA | TCGA-L5-A43M | ASXL2 | chr2 | 26101035 | - | HADHA | chr2 | 26432758 | - |
| ChimerDB4 | UCEC | TCGA-AX-A3FW-01A | ASXL2 | chr2 | 26022254 | - | HADHA | chr2 | 26432758 | - |
Top |
Fusion ORF Analysis |
| Fusion information from ORFfinder translation from full-length transcript sequence from FusionPDB. |
| Henst | Tenst | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand | Seq length (transcript) | BP loci (transcript) | Predicted start (transcript) | Predicted stop (transcript) | Seq length (amino acids) |
| ENST00000435504 | ASXL2 | chr2 | 26101035 | - | ENST00000380649 | HADHA | chr2 | 26432758 | - | 2283 | 351 | 153 | 1667 | 504 |
| ENST00000336112 | ASXL2 | chr2 | 26022254 | - | ENST00000380649 | HADHA | chr2 | 26432758 | - | 2579 | 647 | 551 | 1963 | 470 |
| DeepORF prediction of the coding potential based on the fusion transcript sequence of in-frame fusion genes. DeepORF is a coding potential classifier based on convolutional neural network by comparing the real Ribo-seq data. If the no-coding score < 0.5 and coding score > 0.5, then the in-frame fusion transcript is predicted as being likely translated. |
| Henst | Tenst | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand | No-coding score | Coding score |
| ENST00000435504 | ENST00000380649 | ASXL2 | chr2 | 26101035 | - | HADHA | chr2 | 26432758 | - | 0.001720573 | 0.99827945 |
| ENST00000336112 | ENST00000380649 | ASXL2 | chr2 | 26022254 | - | HADHA | chr2 | 26432758 | - | 0.001844894 | 0.99815506 |
Top |
Fusion Amino Acid Sequences |
| For individual full-length fusion transcript sequence from FusionPDB, we ran ORFfinder and chose the longest ORF among the all predicted ones. |
| >FusionGDB ID_FusionGDB isoform ID_FGname_Hgene_Hchr_Hbp_Henst_Tgene_Tchr_Tbp_Tenst_length(fusion AA) seq_BP >7307_7307_1_ASXL2-HADHA_ASXL2_chr2_26022254_ENST00000336112_HADHA_chr2_26432758_ENST00000380649_length(amino acids)=470AA_BP=32 MVSQIPRVLRTAAAAVMVAATRREKRAGGKGKKFGELVMTKESKALMGLYHGQVLCKKNKFGAPQKDVKHLAILGAGLMGAGIAQVSVDK GLKTILKDATLTALDRGQQQVFKGLNDKVKKKALTSFERDSIFSNLTGQLDYQGFEKADMVIEAVFEDLSLKHRVLKEVEAVIPDHCIFA SNTSALPISEIAAVSKRPEKVIGMHYFSPVDKMQLLEIITTEKTSKDTSASAVAVGLKQGKVIIVVKDGPGFYTTRCLAPMMSEVIRILQ EGVDPKKLDSLTTSFGFPVGAATLVDEVGVDVAKHVAEDLGKVFGERFGGGNPELLTQMVSKGFLGRKSGKGFYIYQEGVKRKDLNSDMD SILASLKLPPKSEVSSDEDIQFRLVTRFVNEAVMCLQEGILATPAEGDIGAVFGLGFPPCLGGPFRFVDLYGAQKIVDRLKKYEAAYGKQ -------------------------------------------------------------- >7307_7307_2_ASXL2-HADHA_ASXL2_chr2_26101035_ENST00000435504_HADHA_chr2_26432758_ENST00000380649_length(amino acids)=504AA_BP=66 MLGCPPGLTSLFSHLSRHRGKSSAGQSRQGGRPEPGHGSLPVSSRPDMREKGRRKKGRTWAEAAKTKFGELVMTKESKALMGLYHGQVLC KKNKFGAPQKDVKHLAILGAGLMGAGIAQVSVDKGLKTILKDATLTALDRGQQQVFKGLNDKVKKKALTSFERDSIFSNLTGQLDYQGFE KADMVIEAVFEDLSLKHRVLKEVEAVIPDHCIFASNTSALPISEIAAVSKRPEKVIGMHYFSPVDKMQLLEIITTEKTSKDTSASAVAVG LKQGKVIIVVKDGPGFYTTRCLAPMMSEVIRILQEGVDPKKLDSLTTSFGFPVGAATLVDEVGVDVAKHVAEDLGKVFGERFGGGNPELL TQMVSKGFLGRKSGKGFYIYQEGVKRKDLNSDMDSILASLKLPPKSEVSSDEDIQFRLVTRFVNEAVMCLQEGILATPAEGDIGAVFGLG -------------------------------------------------------------- |
Top |
Fusion Protein Functional Features |
| Four levels of functional features of fusion genes Go to FGviewer search page for the most frequent breakpoint (https://ccsmweb.uth.edu/FGviewer/chr2:26101035/chr2:26432758) - FGviewer provides the online visualization of the retention search of the protein functional features across DNA, RNA, protein, and pathological levels. - How to search 1. Put your fusion gene symbol. 2. Press the tab key until there will be shown the breakpoint information filled. 4. Go down and press 'Search' tab twice. 4. Go down to have the hyperlink of the search result. 5. Click the hyperlink. 6. See the FGviewer result for your fusion gene. |
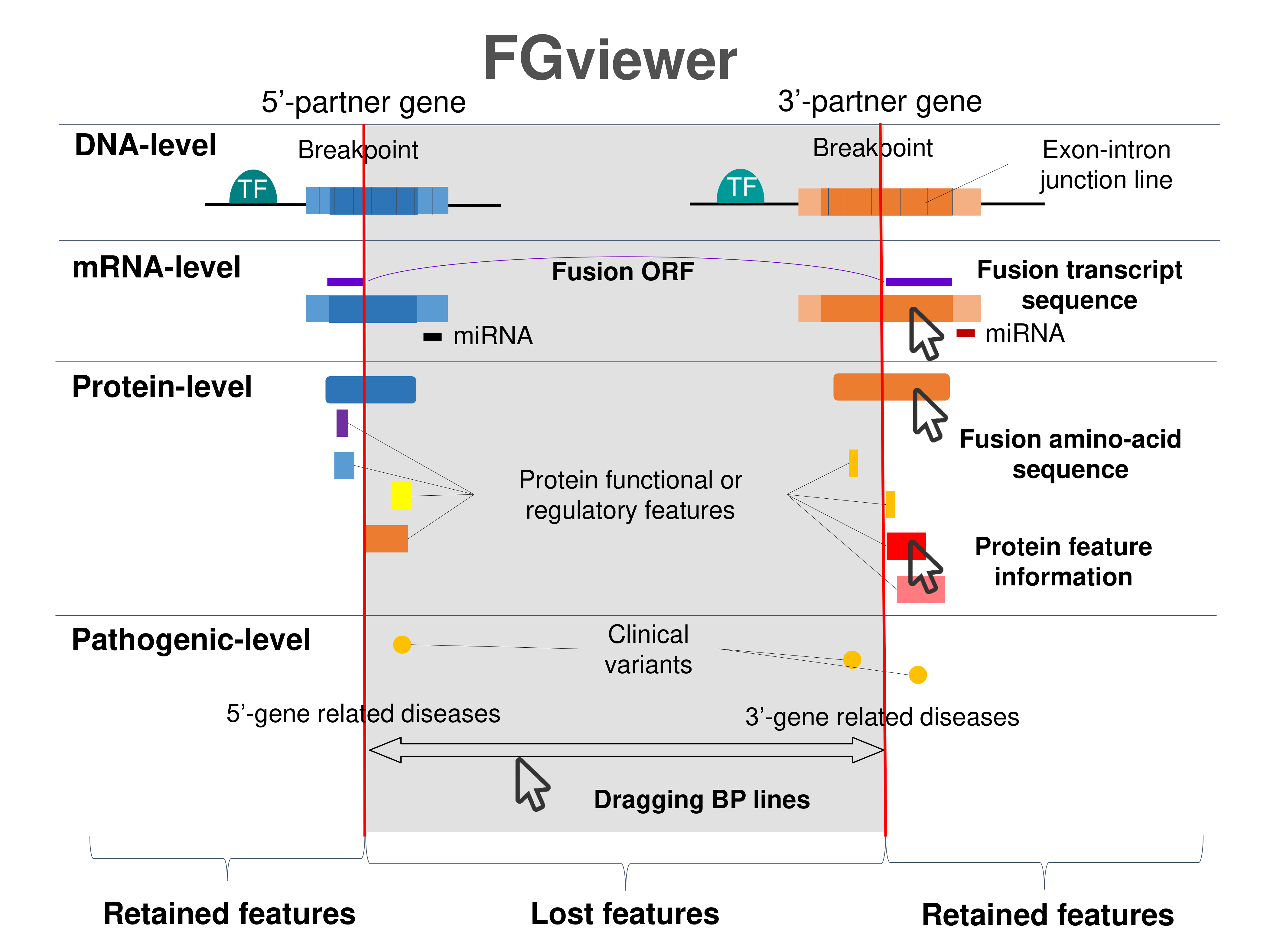 |
| Main function of each fusion partner protein. (from UniProt) |
| Hgene | Tgene |
| ASXL2 | HADHA |
| FUNCTION: Putative Polycomb group (PcG) protein. PcG proteins act by forming multiprotein complexes, which are required to maintain the transcriptionally repressive state of homeotic genes throughout development. PcG proteins are not required to initiate repression, but to maintain it during later stages of development. They probably act via methylation of histones, rendering chromatin heritably changed in its expressibility (By similarity). Involved in transcriptional regulation mediated by ligand-bound nuclear hormone receptors, such as peroxisome proliferator-activated receptor gamma (PPARG). Acts as coactivator for PPARG and enhances its adipocyte differentiation-inducing activity; the function seems to involve differential recruitment of acetylated and methylated histone H3. {ECO:0000250, ECO:0000269|PubMed:21047783}. | FUNCTION: Mitochondrial trifunctional enzyme catalyzes the last three of the four reactions of the mitochondrial beta-oxidation pathway (PubMed:8135828, PubMed:1550553, PubMed:29915090, PubMed:30850536). The mitochondrial beta-oxidation pathway is the major energy-producing process in tissues and is performed through four consecutive reactions breaking down fatty acids into acetyl-CoA (PubMed:29915090). Among the enzymes involved in this pathway, the trifunctional enzyme exhibits specificity for long-chain fatty acids (PubMed:30850536). Mitochondrial trifunctional enzyme is a heterotetrameric complex composed of two proteins, the trifunctional enzyme subunit alpha/HADHA described here carries the 2,3-enoyl-CoA hydratase and the 3-hydroxyacyl-CoA dehydrogenase activities while the trifunctional enzyme subunit beta/HADHB bears the 3-ketoacyl-CoA thiolase activity (PubMed:8135828, PubMed:29915090, PubMed:30850536). Independently of the subunit beta, the trifunctional enzyme subunit alpha/HADHA also has a monolysocardiolipin acyltransferase activity (PubMed:23152787). It acylates monolysocardiolipin into cardiolipin, a major mitochondrial membrane phospholipid which plays a key role in apoptosis and supports mitochondrial respiratory chain complexes in the generation of ATP (PubMed:23152787). Allows the acylation of monolysocardiolipin with different acyl-CoA substrates including oleoyl-CoA for which it displays the highest activity (PubMed:23152787). {ECO:0000269|PubMed:1550553, ECO:0000269|PubMed:23152787, ECO:0000269|PubMed:29915090, ECO:0000269|PubMed:30850536, ECO:0000269|PubMed:8135828, ECO:0000303|PubMed:29915090, ECO:0000303|PubMed:30850536}. |
| Retention analysis result of each fusion partner protein across 39 protein features of UniProt such as six molecule processing features, 13 region features, four site features, six amino acid modification features, two natural variation features, five experimental info features, and 3 secondary structure features. Here, because of limited space for viewing, we only show the protein feature retention information belong to the 13 regional features. All retention annotation result can be downloaded at * Minus value of BPloci means that the break pointn is located before the CDS. |
| - Retained protein feature among the 13 regional features. |
| Partner | Gene | Hbp | Tbp | ENST | Strand | BPexon | TotalExon | Protein feature loci | *BPloci | TotalLen | Protein feature | Protein feature note |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 11_86 | 134.33333333333334 | 1436.0 | Domain | HTH HARE-type |
| - Not-retained protein feature among the 13 regional features. |
| Partner | Gene | Hbp | Tbp | ENST | Strand | BPexon | TotalExon | Protein feature loci | *BPloci | TotalLen | Protein feature | Protein feature note |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 1173_1176 | 0 | 919.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 303_307 | 0 | 919.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 654_684 | 0 | 919.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 687_739 | 0 | 919.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 95_235 | 0 | 919.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 1173_1176 | 0 | 919.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 303_307 | 0 | 919.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 654_684 | 0 | 919.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 687_739 | 0 | 919.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 95_235 | 0 | 919.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 1173_1176 | 134.33333333333334 | 1436.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 303_307 | 134.33333333333334 | 1436.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 654_684 | 134.33333333333334 | 1436.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 687_739 | 134.33333333333334 | 1436.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 95_235 | 134.33333333333334 | 1436.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 1173_1176 | 0 | 919.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 303_307 | 0 | 919.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 654_684 | 0 | 919.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 687_739 | 0 | 919.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 95_235 | 0 | 919.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 1173_1176 | 0 | 919.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 303_307 | 0 | 919.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 654_684 | 0 | 919.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 687_739 | 0 | 919.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 95_235 | 0 | 919.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 1173_1176 | 19.0 | 1436.0 | Compositional bias | Note=Poly-Ser |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 303_307 | 19.0 | 1436.0 | Compositional bias | Note=Poly-Leu |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 654_684 | 19.0 | 1436.0 | Compositional bias | Note=Ala-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 687_739 | 19.0 | 1436.0 | Compositional bias | Note=Gly-rich |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 95_235 | 19.0 | 1436.0 | Compositional bias | Note=Ser-rich |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 11_86 | 0 | 919.0 | Domain | HTH HARE-type |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 274_383 | 0 | 919.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 11_86 | 0 | 919.0 | Domain | HTH HARE-type |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 274_383 | 0 | 919.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 274_383 | 134.33333333333334 | 1436.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 11_86 | 0 | 919.0 | Domain | HTH HARE-type |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 274_383 | 0 | 919.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 11_86 | 0 | 919.0 | Domain | HTH HARE-type |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 274_383 | 0 | 919.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 11_86 | 19.0 | 1436.0 | Domain | HTH HARE-type |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 274_383 | 19.0 | 1436.0 | Domain | DEUBAD |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 174_178 | 0 | 919.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 887_891 | 0 | 919.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 174_178 | 0 | 919.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 887_891 | 0 | 919.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 174_178 | 134.33333333333334 | 1436.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 887_891 | 134.33333333333334 | 1436.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 174_178 | 0 | 919.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 887_891 | 0 | 919.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 174_178 | 0 | 919.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 887_891 | 0 | 919.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 174_178 | 19.0 | 1436.0 | Motif | Nuclear localization signal |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 887_891 | 19.0 | 1436.0 | Motif | Note=LXXLL motif 2 |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000272341 | - | 4 | 13 | 1397_1434 | 0 | 919.0 | Zinc finger | Note=PHD-type%3B atypical |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 1397_1434 | 0 | 919.0 | Zinc finger | Note=PHD-type%3B atypical |
| Hgene | ASXL2 | chr2:26022254 | chr2:26432758 | ENST00000435504 | - | 5 | 13 | 1397_1434 | 134.33333333333334 | 1436.0 | Zinc finger | Note=PHD-type%3B atypical |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000272341 | - | 1 | 13 | 1397_1434 | 0 | 919.0 | Zinc finger | Note=PHD-type%3B atypical |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000404843 | - | 1 | 10 | 1397_1434 | 0 | 919.0 | Zinc finger | Note=PHD-type%3B atypical |
| Hgene | ASXL2 | chr2:26101035 | chr2:26432758 | ENST00000435504 | - | 1 | 13 | 1397_1434 | 19.0 | 1436.0 | Zinc finger | Note=PHD-type%3B atypical |
Top |
Fusion Protein Structures |
| PDB and CIF files of the predicted fusion proteins * Here we show the 3D structure of the fusion proteins using Mol*. AlphaFold produces a per-residue confidence score (pLDDT) between 0 and 100. Model confidence is shown from the pLDDT values per residue. pLDDT corresponds to the model’s prediction of its score on the local Distance Difference Test. It is a measure of local accuracy (from AlphfaFold website). To color code individual residues, we transformed individual PDB files into CIF format. |
| Fusion protein PDB link (fusion AA seq ID in FusionPDB) | Hgene | Hchr | Hbp | Hstrand | Tgene | Tchr | Tbp | Tstrand | AA seq | Len(AA seq) |
| PDB file >>>1069_ASXL2_26101035_HADHA_26432758_1069_ASXL2_26101035_HADHA_26432758_ranked_0.pdb | ASXL2 | 26022254 | 26101035 | ENST00000380649 | HADHA | chr2 | 26432758 | - | MLGCPPGLTSLFSHLSRHRGKSSAGQSRQGGRPEPGHGSLPVSSRPDMREKGRRKKGRTWAEAAKTKFGELVMTKESKALMGLYHGQVLC KKNKFGAPQKDVKHLAILGAGLMGAGIAQVSVDKGLKTILKDATLTALDRGQQQVFKGLNDKVKKKALTSFERDSIFSNLTGQLDYQGFE KADMVIEAVFEDLSLKHRVLKEVEAVIPDHCIFASNTSALPISEIAAVSKRPEKVIGMHYFSPVDKMQLLEIITTEKTSKDTSASAVAVG LKQGKVIIVVKDGPGFYTTRCLAPMMSEVIRILQEGVDPKKLDSLTTSFGFPVGAATLVDEVGVDVAKHVAEDLGKVFGERFGGGNPELL TQMVSKGFLGRKSGKGFYIYQEGVKRKDLNSDMDSILASLKLPPKSEVSSDEDIQFRLVTRFVNEAVMCLQEGILATPAEGDIGAVFGLG | 504 |
Top |
pLDDT score distribution |
| pLDDT score distribution of the predicted wild-type structures of two partner proteins from AlphaFold2 * AlphaFold produces a per-residue confidence score (pLDDT) between 0 and 100. |
ASXL2_pLDDT.png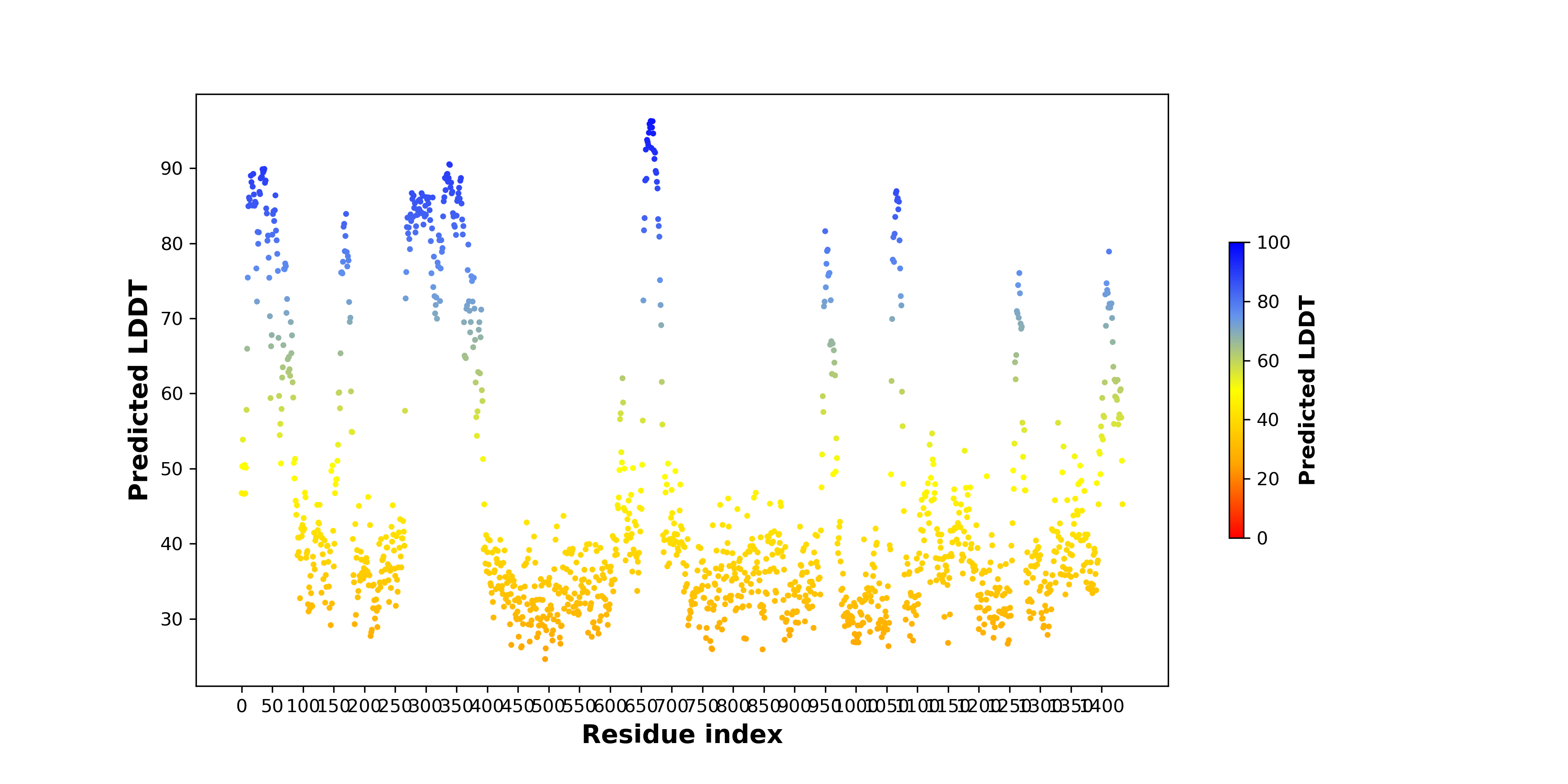  |
HADHA_pLDDT.png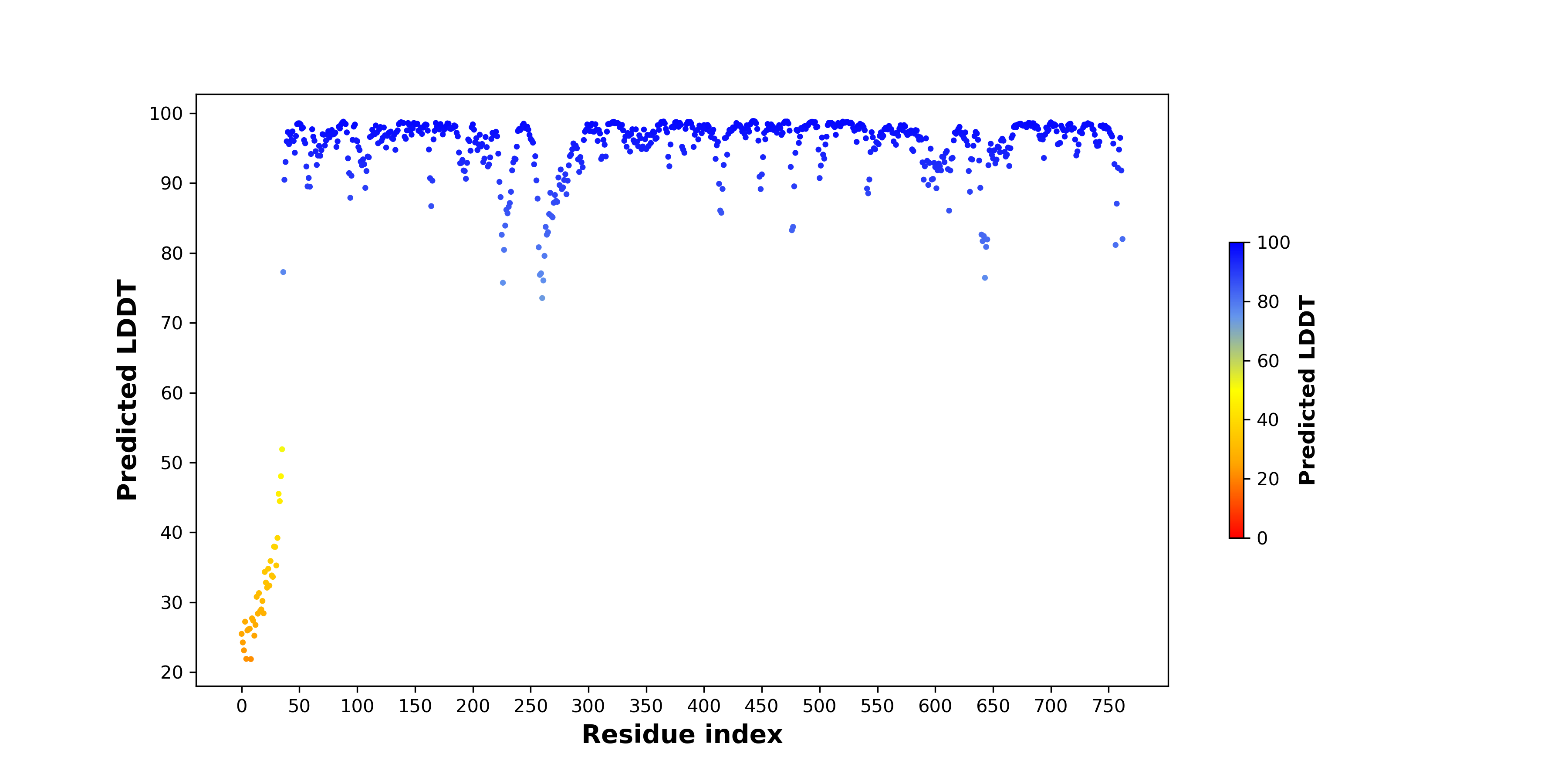  |
| pLDDT score distribution of the predicted fusion protein structures from AlphaFold2 * AlphaFold produces a per-residue confidence score (pLDDT) between 0 and 100. |
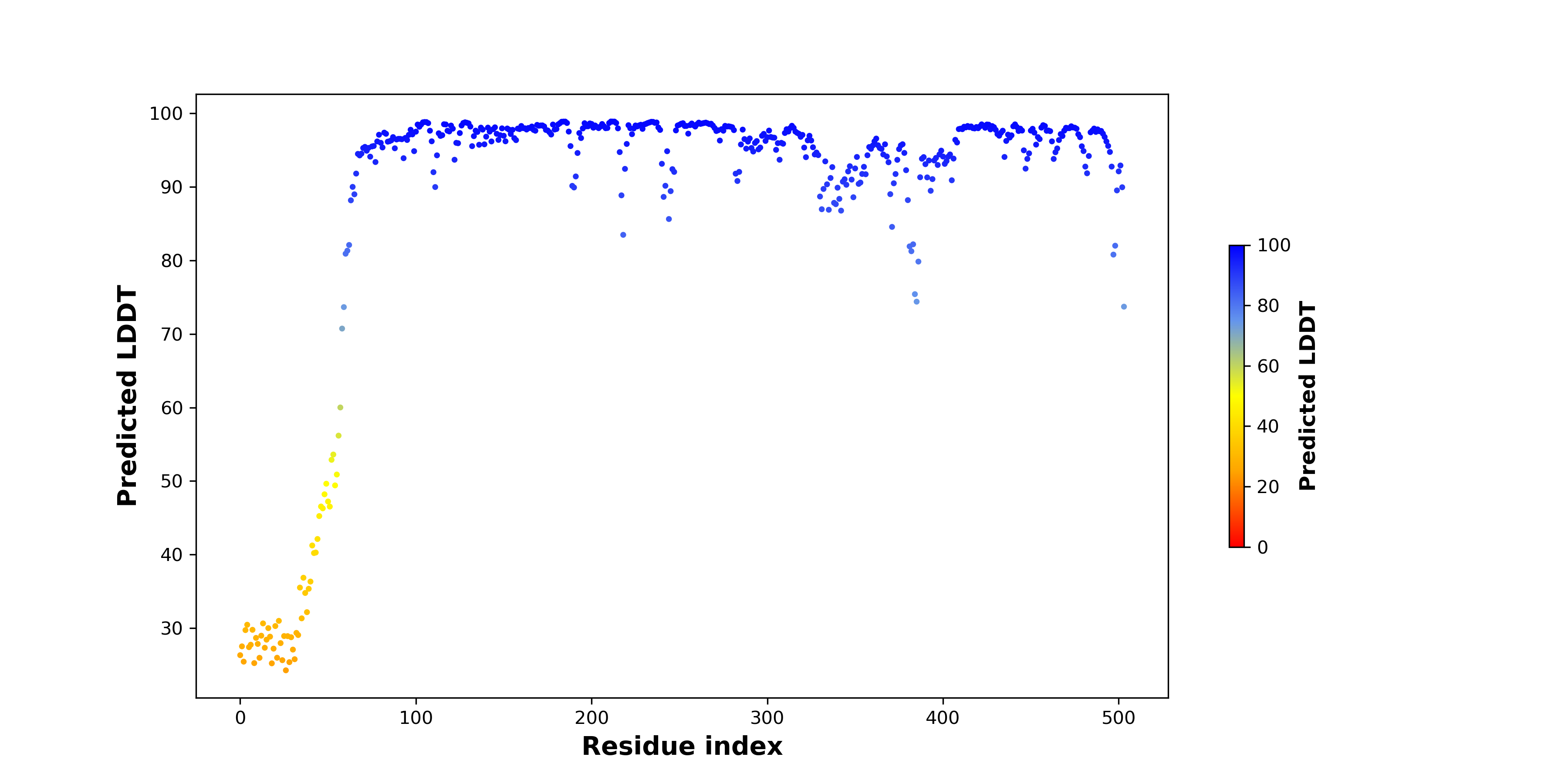 |
Top |
Ramachandran Plot of Fusion Protein Structure |
| Ramachandran plot of the torsional angles - phi (φ)and psi (ψ) - of the residues (amino acids) contained in this fusion protein peptide. |
| Fusion AA seq ID in FusionPDB and their Ramachandran plots |
Top |
Fusion Protein-Protein Interaction |
| Go to ChiPPI (Chimeric Protein-Protein interactions) to see the chimeric PPI interaction in |
| Protein-protein interactors with each fusion partner protein in wild-type from validated records (BIOGRID-3.4.160) |
| Gene | PPI interactors |
| Protein-protein interactors based on sequence similarity (STRING) |
| Gene | STRING network |
| ASXL2 | 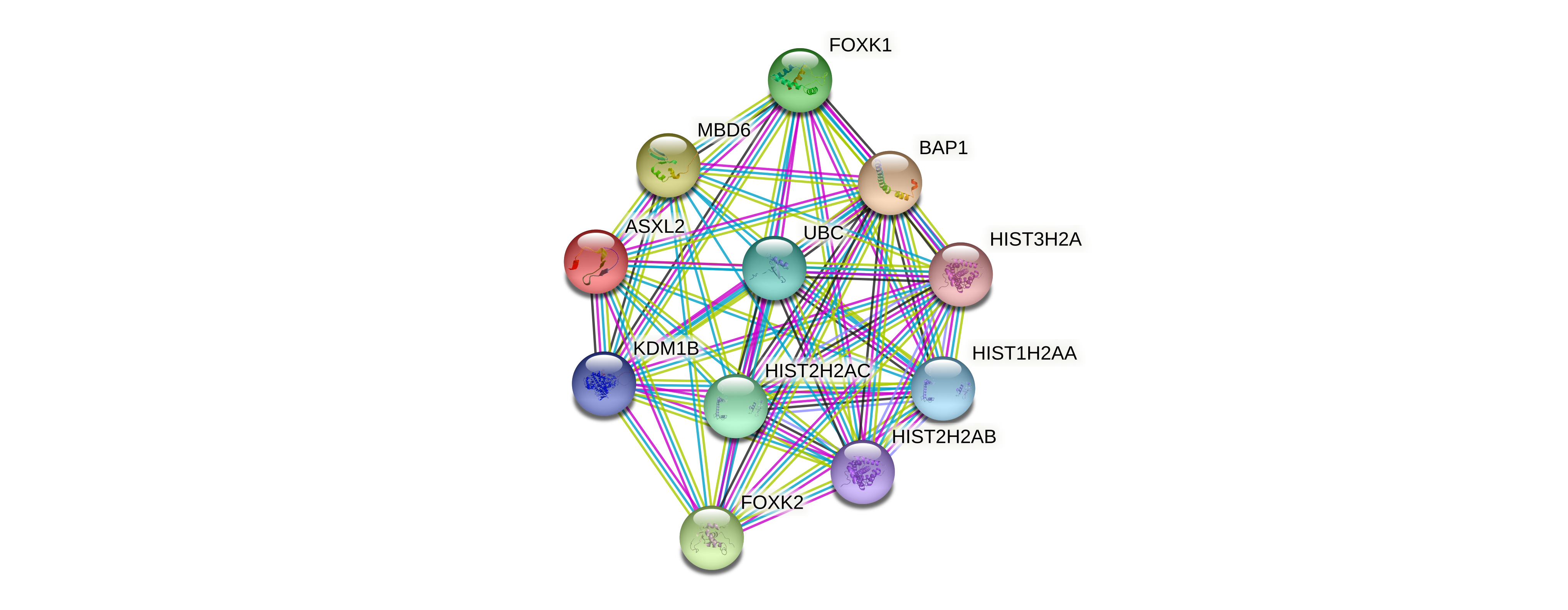 |
| HADHA |
| - Retained interactions in fusion protein (protein functional feature from UniProt). |
| Partner | Gene | Hbp | Tbp | ENST | Strand | BPexon | TotalExon | Protein feature loci | *BPloci | TotalLen | Still interaction with |
| - Lost interactions due to fusion (protein functional feature from UniProt). |
| Partner | Gene | Hbp | Tbp | ENST | Strand | BPexon | TotalExon | Protein feature loci | *BPloci | TotalLen | Interaction lost with |
Top |
Related Drugs to ASXL2-HADHA |
| Drugs used for this fusion-positive patient. (Manual curation of PubMed, 04-30-2022 + MyCancerGenome) |
| Hgene | Tgene | Drug | Source | PMID |
Top |
Related Diseases to ASXL2-HADHA |
| Diseases that have this fusion gene. (Manual curation of PubMed, 04-30-2022 + MyCancerGenome) |
| Hgene | Tgene | Disease | Source | PMID |
| Diseases associated with fusion partners. (DisGeNet 4.0) |
| Partner | Gene | Disease ID | Disease name | # pubmeds | Source |
| Hgene | ASXL2 | C0005684 | Malignant neoplasm of urinary bladder | 1 | CTD_human |
| Hgene | ASXL2 | C0005695 | Bladder Neoplasm | 1 | CTD_human |
| Hgene | ASXL2 | C0023467 | Leukemia, Myelocytic, Acute | 1 | CTD_human |
| Hgene | ASXL2 | C0026998 | Acute Myeloid Leukemia, M1 | 1 | CTD_human |
| Hgene | ASXL2 | C1879321 | Acute Myeloid Leukemia (AML-M2) | 1 | CTD_human |
| Hgene | ASXL2 | C4310672 | SHASHI-PENA SYNDROME | 1 | GENOMICS_ENGLAND |
(UTHealth)
Web File Viewing | Emergency Information |Campus Carry|Site Policies